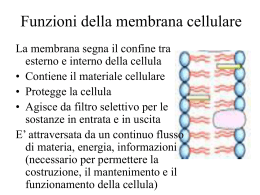
I lipidi di membrana presentano una struttura di base comune a tutti che conferisce loro proprietà complessive analoghe: ANALOGIA Tutti i lipidi sono molecole ANFIPATICHE a causa della coesistenza di parti dotate di proprietà diverse TESTA: è costituita dal gruppo fosfato e dai residui glicidici ed è la sede di cariche elettriche unitarie o parziali; essa costituisce, quindi, nella molecola una regione polare ed idrofila; CODA: è costituita dalle catene idrocarboniose e pertanto è sempre apolare ed idrofoba. Tutto ciò consente ai lipidi di membrana di organizzarsi insieme in una struttura sopramolecolare coerente. La coesistenza di regioni idrofile ed idrofobe ha conseguenze importanti: A. PELLICOLA MONOMOLECOLARE Quando versiamo una preparazione di molecole lipidiche sulla superficie dell’acqua, solvente polare, esse tendono ad assumere una posizione ordinata ponendo le teste idrofile a contatto con l’acqua e rigirando le code idrofobe rivolte verso l’alto fuori dal contatto con l’acqua. A. PELLICOLA MONOMOLECOLARE Questo tipo di disposizione permette al sistema difasico acqua-lipidi di formare al suo interno il maggior numero possibile di interazioni elettrostatiche tra raggruppamenti polari •Acqua-Acqua - Acqua-Teste - Teste-Teste e contemporaneamente il maggior numero di interazioni idrofobiche tra le regioni non polari dei lipidi •Code-Code. Tale disposizione è termodinamicamente favorita perché contribuisce a diminuire il contenuto di energia libera del sistema e viene perciò assunta spontaneamente. La coesistenza di regioni idrofile ed idrofobe ha conseguenze importanti: Formazione di strutture a geometria tridimensionale: B. MICELLE e C. LIPOSOMI. Tali associazioni sopramolecolari si formano nel momento in cui forziamo le molecole lipidiche a penetrare dalla superficie entro la massa d’acqua agitando, per esempio, energicamente il miscuglio acqua-lipidi. B. MICELLE Sono aggregati sferoidali di molecole lipidiche orientate con le teste polari verso la superficie e, quindi, a contatto con l’acqua circostante e con le code idrofobiche disposte tutte verso l’interno dello sferoide a costituire un ambiente interamente idrofobo dal quale le molecole d’acqua sono escluse. C. LIPOSOMI Sono strutture più grandi e più complesse rispetto alle micelle. Sono costituite da un doppio strato di lipidi a disposizione ordinata in cui le teste polari, idrofile, delimitano una superficie esterna ed una interna, mentre le code apolari costituiscono una barriera idrofoba interposta fra le superfici. I liposomi possono così rimanere nell’acqua ed ospitare un contenuto acquoso che rimane escluso dal contatto con l’ambiente esterno. I liposomi costituiscono un modello semplificato, ma plausibile dal punto di vista chimico-fisico, della membrana cellulare eucariotica nonché di quelle intracellulari. LISOSOMI NUCLEO La dimostrazione fu offerta negli anni ‘20 da Gorter e Grendel. Dopo aver estratto i lipidi dalle membrane eritrocitarie, osservarono che versandoli sulla superficie dell’acqua, se ne potevano ottenere foglietti a singolo strato aventi una superficie approssimativamente doppia rispetto a quella degli eritrociti dai quali erano estratti. Ciò indicò, quindi, che nelle membrane eritrocitarie di origine le molecole lipidiche non potevano costituire uno strato semplice, bensì un DOPPIO STRATO. Il foglietto lipidico a doppio strato è una struttura stabile, ma dinamica, dotata di consistenza fluida ed una spiccata asimmetria di composizione che rende le superfici funzionalmente differenti: 1. STABILITA’ 2. FLUIDITA’ 3. ASIMMETRIA 1. STABILITA’ Essa è dovuta alle forze non covalenti che associano tra loro i componenti lipidici: • Interazioni idrofobiche tra le code non polari; • Legami elettrostatici deboli fra le teste polari; • La presenza dell’acqua, su entrambe le superfici del foglietto, che forma con le proprie molecole polari il caratteristico reticolo elettrostaticamente aderente alle teste polari dei lipidi, contribuisce a rendere la disposizione ordinata dei lipidi termodinamicamente vantaggiosa e quindi più stabile. 2. FLUIDITA’ Essa dipende: • Dall’assenza di legami covalenti tra le molecole lipidiche • Dalle caratteristiche degli acidi grassi che entrano nella composizione dei lipidi PROPRIETA’ DISTINTIVE DEGLI ACIDI GRASSI Essi differiscono per: • la lunghezza della catena carboniosa: da 14 a 20 atomi di carbonio; • la presenza (es. acidi grassi insaturi) o l’assenza (es. acidi grassi saturi) nella catena carboniosa di doppi legami. 1) Conformazione di catene lipidiche sature a disposizione rettilinea 2) Conformazione di catene lipidiche insature con angolatura rigida Conseguenze della lunghezza della catena e della presenza di doppi legami sulla fluidità della membrana: •Il numero di interazioni idrofobiche possibile tra le catene brevi sature è minore di quello che può stabilirsi tra le catene lunghe sature e di conseguenza le molecole che si associano mediante interazioni tra catene brevi sature sono meno saldamente unite •I gomiti presenti nelle catene insature contribuiscono ad aumentare lo spazio di separazione tra le molecole contigue Così nei due casi costituiti da catene brevi sature e catene insature, le molecole adiacenti non possono legarsi strettamente le une alle altre DENSO -- DENSO ---DENSO La fluidità della membrana aumenta progressivamente con le catene brevi sature e le catene insature (< Densità ossia > Fluidità) ! Metodi Fisici per la Valutazione della Fluidità della Membrana, un esempio Risonanaza di Spin Elettronico In un foglietto artificiale, lo spostamento di singole molecole lipidiche viene rilevato mediante la coniugazione di raggruppamento chimico contenente un elettrone spaiato. I risultati dimostrano che i lipidi di membrana eseguono: •Rapidi e continui movimenti di rotazione su se stessi •Rapidi e continui movimenti di traslazione sul piano della membrana I risultati dimostrano che: •I lipidi di membrana non eseguono movimenti di capovolgimento che potrebbero portare le molecole ad uscire dallo strato in cui sono contenute per penetrare nello strato opposto del foglietto •L’impulso energetico richiesto perché la testa polare della molecola superi l’ostacolo, costituito dalle code idrocarboniose, è talmente grande che tali movimenti sono praticamente inesistenti! 3. ASIMMETRIA • L’asimmetria del foglietto lipidico è dovuta alla ineguale distribuzione dei diversi tipi di molecole lipidiche tra i due strati del foglietto, ciascuno dei quali ha una sua composizione specifica • Una volta precostituita all’atto della biogenesi, l’asimmetria viene mantenuta definitivamente grazie all’impossibilità dei lipidi di capovolgersi Spazio Extracellulare: fosfatidilcolina, sfingomielina, colesterolo e glicolipidi Versante Citosolico: fosfatidilserina, fosfatidilinositolo, fosfatidiletanolamina e colesterolo COMPONENTE PROTEICA La presenza delle proteine nelle membrane cellulari è chiaramente indicata dai risultati dell’analisi della loro composizione, ma è anche indirettamente suggerita dalla localizzazione, nelle membrane, di attività tipiche delle molecole proteiche quali ad esempio la funzione enzimatica. 1. Quali sono i meccanismi utilizzati per associare le proteine alla membrana? 2. Quali sono le interazioni che le proteine contraggono con il foglietto lipidico? A tale scopo si utilizzano: • Tecniche di tipo estrattivo • Tecniche di visualizzazione Tecniche di Tipo Estrattivo Mediante l’utilizzo delle tecniche di tipo estrattivo ci si propone di separare le proteine dalla membrana e di dedurre il meccanismo della loro associazione al foglietto lipidico dalla natura dei mezzi necessari per separarle Tecniche di Visualizzazione Esse si attuano mediante l’impiego del microscopio elettronico consentendo così di precisare i rapporti spaziali tra proteine e foglietto lipidico. Classificazione delle Proteine di Membrana • Proteine Superficiali o Estrinseche • Proteine Integrali o Intrinseche Proteine Superficiali o Estrinseche • • Questo tipo di proteina ( 1 ) lega la sua superficie a quella formata dalle teste polari giustapposte dei lipidi, oppure alla regione polare di altre molecole inserite tra i lipidi ed affacciate sulle superfici della stessa membrana cellulare. Proteine Superficiali o Estrinseche Esse possono essere separate ed estratte da preparazioni di membrana mediante trattamento con soluzioni saline ad elevata forza ionica al fine di rompere i legami polari non covalenti ovvero le interazioni deboli di natura elettrostatica Proteine Integrali o Intrinseche Le proteine intrinseche (2) sono, come i lipidi di membrana, molecole anfipatiche in grado di legare la loro regione non polare alle code non polari dei lipidi mediante interazioni idrofobiche, mentre le porzioni polari si affacciano verso l’una o l’altra o verso le entrambi le superfici della membrana; esse possono essere, quindi, esposte sia sul versante extra che intracellulare. Proteine Integrali o Intrinseche Esse possono essere separate ed estratte da preparazioni di membrana mediante trattamento con detergenti anfipatici capaci di disgregare completamente il foglietto lipidico della membrana solubilizzandone i componenti. L’azione del detergente è quella, quindi, di annullare le interazioni idrofobiche che contribuiscono alla stabilità della membrana Esempio di Detergenti: SDS e Triton X-100 Q uick Time™ and a TIFF (Unco mpressed ) decompressor are needed to see t his pi cture. • • La regione idrofobica dei detergenti è in verde; La regione idrofilica è segnata, invece, in blu. Modello Strutturale della Membrana Biologica Il Modello del Mosaico Fluido Il Modello del Mosaico Fluido Rappresenta la migliore interpretazione oggi disponibile dell’organizzazione delle membrane a livello molecolare. Il termine mosaico rappresenta l’interposizione di lipidi e proteine stabilizzate interamente o in grandissima parte da interazioni non covalenti. Il Modello del Mosaico Fluido • • • • • Le interazioni idrofobiche garantiscono la solidarietà delle code apolari dei lipidi tra loro e con le porzioni apolari delle proteine integrali Le interazioni polari si stabiliscono fra le teste polari dei lipidi, le porzioni polari delle proteine e l’acqua presente su entrambe le superfici della membrana Al doppio strato lipidico si associano le proteine intrinseche o integrali e quelle estrinseche o superficiali Le proteine intrinseche possono attraversare interamente il foglietto ed esporre parti della loro molecola su una o entrambe le superfici Le proteine estrinseche sono legate alle superfici esclusivamente mediante legami polari sia con le teste dei lipidi, sia con le porzioni superficiali delle proteine integrali Il Modello del Mosaico Fluido Le proteine condividono con i lipidi alcune caratteristiche quali la mobilità e l’asimmetria di distribuzione nei confronti delle superfici della membrana. Mobilità delle Proteine Esperimento di Frye-Edidin, 1970 • Le membrane di cellule di specie diversa sono caratterizzate da proteine diverse • Si possono produrre reagenti immunologici, ossia anticorpi, capaci di riconoscere con grande specificità e precisione le proteine delle due specie diverse • E’ possibile costituire cellule ibride fondendo cellule umane con cellule di topo ed individuare la distribuzione delle proteine umane e murine del prodotto di fusione Mobilità delle Proteine QuickT ime™ and a T IFF (Uncompressed) decompressor are needed to see this picture. Meccanismi che limitano la Mobilità delle Proteine: I DOMINI DI MEMBRANA Quic kTime™ and a TIFF (Uncom press ed) decompressor are needed to s ee thi s pi cture. • • • • Autoassemblaggio; Ancoraggio alle molecole della matrice extracellulare; Ancoraggio alle proteine situate all’interno cellula; Ancoraggio alle proteine situate su un’altra cellula. Presenza delle Barriere di Diffusione: LE GIUNZIONI OCCLUDENTI QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. E’ da notare che • La proteina A deve necessariamente trovarsi sul lato apicale; • La proteina B deve necessariamente trovarsi, invece, sulla superficie laterale e basale della stessa cellula. Asimmetria delle Proteine Essa contribuisce in maniera determinante a diversificare le due superfici di membrana dal punto di vista della composizione e quindi anche da quello funzionale Asimmetria delle Proteine QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. • • • elica singola, eliche multiple oppure manicotto di tipo Presenza di un legame covalente con una molecola lipidica; Interazioni non covalenti con altre proteine di membrana Asimmetria delle Proteine • Proteine con funzione recettoriale espongono il sito di legame sulla faccia della membrana da cui proviene il ligando, ne sono un esempio i recettori ormonali • Le proteine che devono ricavare l’energia necessaria alla loro funzione dall’idrolisi di ATP espongono il sito di legame per l’ATP sul versante intracitoplasmatico della membrana stessa Funzioni delle Proteine nella Membrana Plasmatica Esse possono essere: • Vettori: Pompa di Na+ (Na+/K+) • Connettori: Integrine (actina e matrice cellulare) • Recettori: PDGF (lega il PDGF) • Enzimi: Adenilato Ciclasi (AMP ciclico) Strato Corticale della Cellula Eritrociti Umani QuickTime™ and a TIFF (PackBits) decompressor are needed to see this picture. • • La spectrina instaura connessioni con la membrana tramite le proteine di attacco che la fissano a specifiche proteine transmembrana Essa deve garantire la robustezza meccanica necessaria ai globuli rossi per poter scorrere nei vasi sanguigni QuickTime™ and a TIFF (PackBits) decompressor are needed to see this picture. • • • Ciascun eterodimero di spettrina consiste di 2 catene polipeptidiche antiparallele chiamate e Le estremità dei tetrameri di spettrina sono unite tramite il legame con filamenti di actina e con la proteina della banda 4.1 formando un complesso giunzionale; La proteina responsabile dell’attacco della spettrina al citoscheletro è l’anchirina che si lega da una parte alla spettrina e dall’altra al dominio citoplasmatico della proteina transmembrana della banda 3. Strato Corticale della Cellula Eritrociti Umani QuickT ime ™an d a TIFF (P ackB its) deco mpre ssor are n eede d to s ee this picture . Rete di spectrina al microscopio elettronico Il Glicocalice • • • I lipidi presenti nella membrana plasmatica possono legare catene di zuccheri generando così i glicopilidi; Le proteine della membrana plasmatica possono legare brevi catene di zuccheri, ovvero oligosaccaridi, generando così glicoproteine; Le proteine di membrana possono legare anche una o più catene lunghe di zuccheri generando i cosiddetti proteoglicani. Tutti i carboidrati appartenenti ai glicolipidi, alle glicoproteine ed ai proteoglicani contribuiscono a formare il GLICOCALICE che si affaccia sempre sul versante non citosolico della cellula. Il Glicocalice QuickTi me™ and a TIFF (U ncompressed) decompressor are needed to see this picture. E’ da notare che alla composizione del glicocalice possono contribuire anche le glicoproteine ed i proteoglicani secreti dalla stessa cellula e subito adsorbiti alla sua superficie. Funzioni del Glicocalice • • • Esso aiuta a proteggere la superficie cellulare dal danneggiamento per cause chimiche e meccaniche; Le catene di zuccheri assorbono acqua rendendo così scivolosa la membrana cellulare: questa caratteristica aiuta per esempio le cellule mobili come i leucociti nella migrazione trans-endoteliale ed evita agli eritrociti di aderire tra di loro o alle pareti dei vasi; Il glicocalice svolge un ruolo fondamentale nel riconoscimento e nell’adesione cellulare. Il Glicocalice è quindi un involucro che distingue cellule specializzate in una particolare funzione e le rende riconoscibili ad altre cellule con cui devono interagire. Risposta Infiammatoria La risposta infiammatoria rappresenta il processo attraverso cui i neutrofili vengono reclutati dal sangue in un sito infiammatorio per combattere ad esempio un’infezione: •Mediatori chimici locali inducono l’espressione sulle cellule endotelali di una glicoproteina transmembrana chiamata selectina P; •Le selectine contengono una lectina che lega specifici carboidrati presenti al livello dei glicolipidi e glicoproteine dei neutrofili; •Questo tipo di legame è responsabile del “rolling” cui segue •Un altro meccanismo di rafforzamento dell’adesione mediato da altre proteine transmembrana chiamate integrine; •Migrazione trans-endoteliale dei granulociti neutrofili. Funzioni del Glicocalice Il riconoscimento dei carboidrati di superficie sui granulociti neutrofili è la fase preliminare della loro migrazione fuori dal torrente circolatorio nella sede dell’infezione. QuickTi me™ e un decompressore TIFF (Non compresso) sono necessari per visualizzare quest'i mmag ine. E’ da notare che le cellule endoteliali se da una parte esprimono specifici recettori che sono in grado di legare le selettine ed integrine linfocitarie, dall’altra secernono chemochine che rafforzano i processi di interazione dei linfociti con le cellule endoteliali stesse. Il Trasporto di Membrana La membrana cellulare è una vera e propria barriera. Tuttavia la cellula deve vivere scambiando molecole con l’ambiente. Infatti, una cellula deve importare sostanze nutritizie, come zuccheri ed amminoacidi, eliminare prodotti di rifiuto, come anidride carbonica, e regolare anche la concentrazione intracellulare di ioni come Idrogeno, Sodio, Potassio e Calcio. Il Trasporto di Membrana Le molecole proteiche svolgono un ruolo fondamentale in questo tipo di processo. Si possono distinguere due classi principali di proteine di trasporto: Proteine di Trasporto Proteine Canale Il Trasporto di Membrana QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. • • Le proteine di trasporto o vettrici vanno incontro ad una serie di cambi conformazionali per trasportare molecole idrosolubili al di là del doppio strato lipidico. In particolare, la proteina trasportatrice si alterna fra 2 conformazioni così che il sito di attacco del soluto sia accessibile sequenzialmente su di un lato e sull’altro del doppio strato Le proteine canale o canali ionici formano piccoli pori idrofilici pieni d’acqua attraverso i quali possono diffondere ioni specifici. Il Trasporto di Membrana: Passivo o Attivo QuickTime™ and a TIFF (PackBits) decompressor are needed to see this picture. • • Se i soluti sono privi di carica o liposolubili, l’attraversamento del doppio strato lipidico avviene per DIFFUSIONE SEMPLICE; La maggior parte dei soluti richiede proteine vettrici oppure canali ionici e l’attraversamento in questi casi può essere PASSIVO, assecondando il gradiente di concentrazione, oppure ATTIVO (esclusivo delle proteine vettrici), contro gradiente di concentrazione con apporto energetico. I Componenti del Gradiente Elettrochimico GRADIENTE ELETTROCHIMICO (Forza Motrice Netta)= Gradiente di Concentrazione + Differenza di Potenziale (tra i due lati della membrana) Le membrane plasmatiche hanno una differenza di potenziale elettrico (gradiente di voltaggio) che le attraversa con la parte interna di solito negativa rispetto a a quella esterna Entità del Gradiente Elettrochimico per lo stesso Soluto QuickTime™ and a TIFF (PackBits) decompressor are needed to see this picture. A= si ha soltanto un gradiente di concentrazione B= al gradiente di concentrazione del soluto si aggiunge un potenziale di membrana che accresce la Forza Motrice C= il potenziale di membrana riduce la forza motrice creata dal gradiente di concentrazione favorevole Tre tipi di Trasporto Passivo o Attivo QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. • • • Uniporto: il trasportatore traghetta un solo tipo di soluto; Simporto: il trasportatore traghetta due soluti nella stessa direzione; Antiporto: il trasportatore traghetta due soluti in direzioni opposte. Il Trasporto di Membrana: Le proteine Vettrici • • • Le proteine vettrici sono altamente selettive Il sito di legame delle proteine vettrici per il soluto che devono trasportare è assolutamente specifico Le proteine vettrici si diversificano a seconda delle membrane cellulari cui sono associate Le proteine Vettrici nel Trasporto Passivo QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. La proteina trasportatrice può esistere in 2 stati conformazionali: • Nello stato A i siti di legame sono esposti all’esterno del doppio strato e • viceversa nello stato B. Sufficienti prove sperimentali indicano che la transizione fra i due stati avvenga a caso e che sia completamente reversibile Le proteine Vettrici: un esempio di Trasporto Passivo Uniporto QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. Il Trasporto Netto del Soluto è secondo Gradiente di Concentrazione Trasporto Passivo del Glucosio al livello dell’Epatocita > Conc. Glucosio all’esterno (dopo un pasto)=Transiz. da A a B Conc. Glucosio all’interno (a digiuno)=Transiz. da B ad A Le proteine Vettrici: il Trasporto Attivo QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. Il trasporto attivo contro gradiente elettrochimico riveste un ruolo fondamentale nel mantenimento della composizione ionica intracellulare e per importare soluti maggiormente concentrati all’interno della cellula rispetto all’esterno. Il Trasporto Attivo permette lo spostamento dei soluti contro il loro gradiente elettrochimico ed avviene in tre modi. A:Le Pompe ad ATP accoppiano il trasporto contro gradiente all’idrolisi di ATP B:I Trasportatori Accoppiati accoppiano il trasporto di un soluto contro gradiente al trasporto di un soluto secondo gradiente C: Le Pompe Fotoalimentate, presenti nei batteri, accoppiano il trasporto contro gradiente all’assorbimento di energia luminosa Trasporto Attivo Antiporto: Le pompe Sodio-Potassio ad ATP QuickTime™ and a TIFF (Uncompressed) decompressor are needed to see this picture. Questa proteina vettrice usa l’energia di idrolisi dell’ATP per espellere Sodio fuori dalla cellula ed importare Potassio dentro la cellula, entrambi contro gradiente di concentrazione. Trasporto Attivo: Le pompe Sodio-Potassio ad ATP QuickTim e™ and a TIFF (Uncom pressed) decom pressor are needed to see this picture. • • • Il legame di Sodio e la successiva fosforilazione inducono il cambio conformazionale e l’espulsione di Sodio stesso; Il legame del Potassio e la successiva defosforilazione riportano da un lato la proteina al suo stato conformazionale originario e dall’altro introducono Potassio all’interno della cellula; Il risultato netto di un ciclo operativo è il trasporto di tre di Sodio e due di Potassio. Trasporto Attivo: Il Trasportatore Accoppiato • L’accoppiamento può avvenire tra movimenti di due ioni organici, tra il movimento di uno ione inorganico e quello di una molecola organica o tra movimenti di due molecole organiche; • I tipi di trasporto possono essere antiporto e simporto. Trasporto Attivo: Il Trasportatore Accoppiato • • Il principio base prevede che se manca uno dei soluti cotrasportati, il trasporto dell’altro non può avvenire; Se le cellule dell’epitelio intestinale disponessero solo dei vettori passivi per il glucosio, lo assumerebbero dopo un pasto ricco di zuccheri e lo libererebbero dopo un pasto privo di zuccheri. Trasporto Attivo: Il Trasportatore Accoppiato nelle Cellule dell’Epitelio Intestinale In realtà l’enterocita può assumere attivamente glucosio anche quando la sua concentrazione intracellulare supera quella dell’intestino grazie alla presenza di un vettore simporto, glucosio-Na+, presente sulla superficie apicale dell’enterocita stesso QuickTi me™ and a TIFF (U ncompressed) decompressor are needed to see this pi cture. Il processo porta al trasporto di glucosio dal lume intestinale al fluido extracellulare ed infine al sangue: • Il glucosio è importato al livello apicale da un simporto di glucosio spint dal Na+ • Il glucosio passa fuori dalla cellula per diffusione facilitata mediata da una proteina diversa al livello dei domini laterali e basali della cellula; • Il gradiente di Na+ che sospinge il simporto di glucosio è mantenuto dalle ATPasi dei domini basali che a loro volta abbassano la concentrazione interna di Na+ stesso Il Trasportatore Accoppiato Glucosio-Na+ Se però l’epitelio intestinale avesse solo il simporto, non potrebbe mai rendere disponibile il glucosio alle altre cellule del corpo. Il Trasportatore Accoppiato Glucosio-Na+ QuickTi me™ and a TIFF (U ncompressed) decompressor are needed to see this pi cture. Infatti, le cellule dell’epitelio intestinale posseggono nei domini laterali e basali il sistema uniporto passivo che liberano glucosio secondo gradiente. E’ da notare la presenza di una barriera antidiffusione assicurata dalle giunzioni occludenti intorno alle regioni apicali. OSMOSI Se la concentrazione dei soluti dentro la cellula è più alta rispetto a quella dell’ambiente extracellulare, l’acqua entra per osmosi. Se il dislivello di concentrazione dei soluti è abbastanza forte, la cellula scoppia.
Scarica